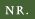| Paljudes hermafrodiitsetes õites hoiduvad isas- ja emaspool teineteisest eemale nii nagu selles päevaliilias.
| Jaan Liira |
Kui õies on tolmukad ja emakad, siis ei pea neis olevad sugurakud tingimata omavahel kokku saama ning ühinema. Nii kaotaks taim palju võimalusi. Looduslikes tingimustes on kasulikum võtta seksist, mis võtta annab. Vastasel juhul ei oleks mingit mõtet raisata materjali ja energiat kahe soo ja sugurakkude peale, kaunitest värvidest, uhketest kroonidest, hurmavatest lõhnadest ja magusatest nektaritest rääkimata.
Loomariigis, kust pärineb ka inimene, on isaseks ja emaseks olemine norm. Seepärast on vahel raske pidada kahesugulisust ehk hermafrodiitsust normaalseks. Aga taimeriigis on see just nii: hinnanguliselt kuulub vähemalt 70% õistaimeliikidest mitte ainult hermafrodiitide hulka, vaid neil on sealjuures ka kahesugulised õied.
Enamik neist kaunitest õitest, mis meid ümbritsevad, sisaldab nii isas- kui ka emasosa tolmukaid ja üht või mitut emakat. Taimedest rääkides on ajapikku isegi tavaks saanud, et just selliseid õisi nimetatakse täielikeks ja ühesugulisi mittetäielikeks.
Üks enim levinud väärarvamusi hermafrodiitsuse puhul seostub valdava isetolmlemisega: arvatakse, et kui õies on tolmukad ja emakad, siis saavad neis olevad sugurakud ka vältimatult kokku ning ühinevad. Kui see oleks tõesti nii, siis kaotaks suguline protsess suure osa oma võimalustest: kahe vanema pärilikku informatsiooni kandvate sugurakkude kombineerumisel saadav mitmekesisus oleks piiratud ainult ühe vanema ja tema geneetilise informatsiooni kombinatsioonidega.
Tomat, nisu ja teised tuntud isetegijad. Isetolmlejaid taimi teatakse palju, tuntuimad neist on kultuurtaimed: suur osa nii tomati, nisu, kaera, odra kui ka aedoa sortidest ei vaja viljumiseks teise taime õietolmu. Klassikalise isetolmleja näiteks sobib harilik hernes. Samas ei lase end sordiaretuselgi isetolmlejaks painutada ei rukis ega mais.
Isetolmlemine, nagu ka sugutu paljunemine sobib juhul, kui on vaja taastoota juba välja töötatud head mudelit. Muutuvates tingimustes, kus kord on eelis ühel, kord teisel variandil, ei osutu isetolmlemine enamasti kasulikuks. Kultuurtaimede puhul on inimene valinud tootlikud sordid, mis tema enda kujundatud keskkonnatingimustes end õigustavad, kuid looduses konkureerida ei suuda. Osa isetolmlejaid viljastub ainult oma õietolmuga, tolmeldamine teiselt taimelt pärineva õietolmuga ei anna tulemusi või õnnestub vaid tühisel määral. Näiteks aedoa katselisel risttolmeldamisel arenesid viljad vaid kuni ühel protsendil õitest.
Iseviljastamine on tavaline ka looduses: rohkem kui pooled parasvöötme õistaimeliikidest võivad viljuda isetolmlemise järel. Mõnel juhul on see viimane väljapääs. Parem ise kui ilma annab suurema hulga geneetiliselt sarnaseid järglasi. Kui need järglased antud tingimustesse sobivad, siis ei olegi tingimata vaja varieerida. Isetolmlemine annab viljastusvõimaluse ka piirkondades, kus tolmeldajaid napib kõrgel mägedes ja kaugel põhjas.
Imekannikese ime ja jänesekapsa saladus. On liike, mis võivad viljastuda nii võõrast kui ka endast. Sellistes õites luuakse kõigepealt eeldused risttolmlemiseks: õitsemise alguses pöörduvad emakad ja tolmukad üksteisest eemale. Õitsemise lõpu poole, kui on karta, et oodatud kauged kosilased õigel ajal kohale ei jõua, lähenetakse teineteist taas leides uuesti: vananevad emakasuudmed painduvad tolmukate poole või tolmukad keerduvad emakasuudme kohale.
Imelike isetolmlemist soodustavate juhtumite hulka võiks arvata väikese hiiresaba, kes saab end ise tolmeldada vaid õitsemise alguses. Hiljem, kui õiepõhi pikeneb, tõstes emakkonna tolmukatest kõrgemale, see enam nii lihtsalt ei käi.
Looduslikel taimedel, kel isetolmlemine ei ole varuväljapääs, vaid tavaline paljunemisviis, on muutunud ka õite välimus: risttolmlejatest sugulasliikidega võrreldes on nad väiksemad ja tagasihoidlikuma õiekattega ning tolmukapead painduvad emakasuudmete kohale. Et võõraist hoidumine oleks kindlam, käib isetolmeldamine veel avanemata õies. Nii teeb näiteks meil haruldane vesilobeelia.
Iseviljastumine on veel tugevamini kinnistunud umbsigiste (kleistogaamsete) õite puhul, mille näiteks sobib imekannike. Ime selles seisnebki, et kevadel õitseb ta nagu ilus siniste õitega kannike ikka, aga vilju neist õitest sageli ei tule. Hiljem arenevad uued õiepungad, mis nagu jääksidki pungadeks. Need kleistogaamsed õied ei uhkelda värvilise krooniga ega avane. Viljastamine toimub mõne otse emakasuudme vastu puutuvast tolmukotist pärit tolmuteraga, valmivad idanemisvõimelised seemned. Ebasoodsates tingimustes tekitavad umbsigiseid õisi teisedki kannikeseliigid, näiteks Eestis haruldane kõrge kannike. Selliseid õisi oskab moodustada ka harilik jänesekapsas: peale meile tuntud valgete lillaka soonestusega õite tekivad tal hiljem maad ligi hoiduvad ja mitte kunagi avanevad väikesed pungõied. Suletud isetolmlevaid õisi moodustab veel aiaumbrohi hõlmine iminõges.
Enamik esmapilgul hermafrodiitseid taimi on evolutsiooni jooksul välja töötanud hulga kavalusi vältimaks iseviljastumist.
Erikaelsus ehk kuidas omadest hoiduda. Erikaelsuse (heterostüülia) puhul on tolmukad ja emakad eri pikkusega. Nii takistatakse oma õietolmu sattumist emakasuudmele. Kui emakakael küünib kõrgele üle tolmukate, siis on ilmselge, et oma õietolm emakasuudmele naljalt ei kuku. Näiteks nurmenuku õide sisse vaadates paistab ühel juhul emakasuudme nupp, teisel juhul varjavad sissepääsu viis krooniputke tolmukapead. Teada on, et äärmuslikes tingimustes kõrgmägedes ja kaugel põhjas kasvavatel nurmenuku perekonna liikidel on isetolmlemist takistav erikaelsus vähem levinud. Ka siis, kui tolmukad on emakakaelast kõrgemal, takistavad iseviljastamist sageli tolmuterade ja neid vastu võtva emakasuudme papillide suuruse sobimatus. Ühe ja sama liigi eri tüüpi õisi ja nende bioloogilist mõtet käsitles põhjalikult juba Charles Darwin oma töödes [3].
Enne meheks, siis naiseks, või vastupidi. Ka taimeriigis on omad transseksuaalid. Järjestikuse ühesugulisuse (dihhogaamia) puhul ei valmi tolmukad ja emakad samal ajal. Sootunnuste ajaline nihe on tuntud eelisasuse (proterandria) ja eelemasusena (proterogüünia): isas- või emasorganid valmivad varem ja minetavad oma talitlusvõime vastassoo küpsuse ajaks. Taimi, mille õies tolmukate suguküpsuse aegu emakast veel asja pole, on meiegi flooras. Näiteks ahtalehine põdrakanep ja harilik luigelill, aga ka mündid ja kellukad, paljud korvõielised ning sarikalised. Lemmaltsa ja kurereha perekonnas kuivavad ja varisevad tolmukad enne, kui õie emasosa valmib. Heites pilgu selles staadiumis olevasse õide, võib ekslikult arvata, et õis on juba esialgu ühesuguline. Niisiis talitleb üks ja sama hermafrodiitne õis alguses isasena ja seejärel emasena.
Ka vastupidi talitlevaid, st. esmalt emaseid õisi leiab hõlpsasti. Kas või teelehtedel, kelle õisikus on emasstaadiumis õied isastena talitlevatest ülalpool, nii et pelgalt raskusjõu mõjul varisev õietolm oma õite emakasuudmeile ei satu. Eelemasust on leitud ka kuuskjala perekonnas, kuid huuljate õite sisemusse peidetud tolmukad ja emakad ei torka siin silma.
Isesobimatus isegi sama kloon pole soositud. Isesobimatus tähendab, et taim ei viljastu oma õietolmuga. Isesobimatuid on nii tuul- kui ka putuktolmlejate seas, kuid just esimeste puhul on oluline tunda emakasuudmel ära õige tolmutera, kuna putuktolmlejate õie iseärasuste ja tolmeldajate täpsem töö sageli juba välistab õietolmu sattumise sama õie emakasuudmele. Samas peab tolmutera ja emakasuudme vahel olema valikuline filter, mis ühelt poolt soodustab sama liigi õietolmu idanemist ja teiselt poolt takistab mitte ainult sama õie tolmuga viljastamist (autogaamia), vaid ka sama taime teise õie või sama klooni õietolmuga viljastamist (geitonogaamia). Viimane tagab populatsiooni geneetilise varieeruvuse just vegetatiivselt hästi paljunevate ja seemneliselt harva uuenevate taimeliikide puhul, sest ükski mehhaanilise ja ajalise isolatsiooni mehhanism, nagu ka tolmeldajate liigispetsiifilisus geitonogaamiat ei välista.
Kuidas tunneb emakasuue ära ja laseb ligi vaid soovitu? Õistaimede emakasuue on loodud tolmuterasid püüdma, kinni hoidma ja idandama. Emakasuudme hõlmade suurus ja kuju, näsade olemasolu jms. on kooskõlas selle õietolmuga, mille vastu võtmiseks ta on evolutsiooniliselt kujunenud. Liigispetsiifilisus ja emaka valiv osa ei ole siin suur, ka tolmutera kuju ja pinnamustri erinevused ei ole enamasti sedavõrd määravad, et takistaks liigile sobimatu tolmu püsimajäämist emakasuudmele, ammugi ei välista ainuüksi see iseviljastamist.
Sobiv tolmutera küpsele emakasuudmele. Tolmutera vajab idanemiseks eelkõige vett ja hapnikku ning idaneb seal, kus tingimused soodsad. On juhtunud, et tolmuterad idanevad nii niiskel kroonlehel [4] kui ka nektaariumil [13]. Sellist tolmutera idanemist väljaspool emakat võiks mõnel juhul nimetada ka lihtsalt lõhkemiseks. Tolmutoru kasvuks peab olema sobiva osmootse rõhuga lahus ja pidev süsivesikute allikas.
Emakasuue on vastuvõtuvõimeline lühikest aega. Kui sobiv tolmutera satub küpsele emakasuudmele, kinnitub ta sinna. Üks silmaga nähtav emakasuudme valmisoleku tunnus on suuet katvate näsakeste turgor ehk prinkolek. Emakasuudmed jagunevad märgadeks (kleepuvateks) ja kuivadeks, kusjuures emakasuudme tüüp on tihedalt seotud isesobimatuse tüübiga [13].
Märjale emakasuudmele kinnitub (kleepub) tolmutera hõlpsasti, samas on tagatud ka tolmutera idanemiseks ja kasvuks sobivad esmased tingimused, emakasuudme näsakesed ei kuiva.
Kuivale emakasuudmele kinnituvad tolmuterad oma välise kihi abil: nad lahustavad emakasuudme pindmist kihti keemiliselt ja saavad seejärel idanemiseks vajaliku vee kätte näsarakkudest. Märjale emakasuudmele kohanenud tolmutera ei ole enamasti võimeline idanema kuival emakasuudmel.
Tolmutoru ei jõua alati pärale. Igast idanenud tolmuterast tekib tolmutoru, mis tungib emakakaela rakkude vahele. Kuidas just, see sõltub emakakaela tüübist. Emakakael võib olla kas umbne, täidetud erilise koega (kaheidulehelistel) või tolmutoru jaoks ettenähtud kanaliga (üheidulehelistel). Tolmutoru kasvu kiirus oleneb temperatuurist, sobivaim on suurel osal liikidest 1040 °C. Edasinihkumise kiirus 1,7535 mm/h ei olene tolmutera suurusest. Tee pikkus erineb liigiti, küündides 1200 mm. Valdaval osal õistaimedest jõuab tolmutoru seemnealgmeni 1248 tunniga [13]. Tolmutoru kasvul on täheldatud tugevat kemotropismi: kui üks tolmutoru on jõudnud seemnealgmeni, peatatakse teiste kasv.
Liigi piires on tolmutoru pikkus ja emakakaela pikkus kooskõlas. Pika emakakaelaga liikide tolmeldamine lühikese emakakaelaga liikide tolmuga võib lõppeda ebaõnnestumisega (1. joonis, c), vanemliikide vastupidise ristamise korral võib tolmeldamine õnnestuda [2].
Soovimatu tolmutoru areng võidakse peatada emakakaela eri piirkondades.
Teatud olukordades (1. joonis, a) ei hakka tolmutoru üldse kasvama. See juhtub nii kaugete liikide ristamisel kui ka sporofüütse isesobimatuse tõttu [2], sellest lähemalt edaspidi. Tolmutoru kasv lakkab eri arengufaasides (1. joonis, bd) nii füsioloogilis-biokeemilist laadi põhjustel (tolmutoru ei saa piisavalt toitaineid normaalseks ainevahetuseks) kui ka sobimatuse tõttu. Kaugristamise puhul tolmutoru kasv tavaliselt aeglustub ja lakkab varsti pärast idanemist (1. joonis, b). Tolmutorud, mille kasv on peatatud, võivad haruneda ja tipus puhetuda. Neid võidakse sundida peatuma või isegi otsa ringi pöörama veel ka üsna kaugele jõudnuna (1. joonis, d).
Isesobimatud ei saa ise hakkama. Isesobimatus (self-incompatibility) on hermafrodiitse õistaime võimetus isetolmlemise abil sügoote moodustada. See võib väljenduda mitmeti: oma õietolm ei kinnitu emakasuudmele, ei idane oma emakasuudmel, tolmutoru ei kasva läbi emakasuudme või emakakaelas (1. joonis). Kõiki neid nähtusi põhjustab geneetiliselt nn. multialleelne isesobimatus (multi-allelic self-incompatibility), mille määravad S-lookuse alleelsete geenide kombinatsioonid. Eristatakse kahte tüüpi multialleelset isesobimatust: gametofüütset (2. joonis) ja sporofüütset.
Gametofüütset isesobimatust on kõige enam uuritud maavitsalistel. Nüüdseks on lisandunud teavet isesobimatuse kohta ka magunalistel, roosõielistel ja mailaselistel [8]. Nendes sugukondades kontrollib gametofüütset isesobimatust üks multialleelne S-lookus, mis vastutab emakakaela eriliste glükoproteiinide eest. Viimased tungivad isesobimatutesse tolmutorudesse ja lagundavad neis kasvuks vajaliku ribosoomi-RNA [9, 10]. Selle tõttu peatub tolmutoru kasv emakakaela ülemises kolmandikus (2. joonis).
Gametofüütne isesobimatus on oma nimetuse saanud sellest, et isesobimatuse reaktsioon sõltub siin tolmutera (gametofüüdi) haploidses genotüübis sisalduva S-alleeli ja emaka diploidses genotüübis sisalduva sama S-alleeli kontrollitud valguliste produktide koostoimest (2. joonis). Gametofüütse isesobimatuse puhul on S-alleelid valdavalt kodominantsed.
Suurem osa isesobimatuid õistaimi on gametofüütselt isesobimatud ja seda süsteemi peetakse ka algelisemaks [13]. Ühe sugukonna piires on leitud ainult kas gametofüütset või sporofüütset isesobimatust.
Sporofüütse isesobimatuse uurimine on keskendunud ristõieliste sugukonnale, mille mõnede liikide isesobimatus on praeguseks molekulaarsel tasemel tõestatud [11]. Nii nagu gametofüütse isesobimatuse puhul, on siingi tegemist multialleelse S-lookusega [8], kuid selle struktuur ja valgulised produktid on teistsugused, kui gametofüütse isesobimatuse puhul. Täpne toimemehhanism tolmutera kasvu peatamisel ei ole seni üheselt selge, kuid selle tõttu peatub tolmutera idanemine juba emakasuudme pinnal.
Sporofüütseks isesobimatuseks nimetatakse seda seetõttu, et reaktsioon sõltub siin tolmutera vanema, tema diploidse emaraku (arhespoori) genotüübist. Erinevalt gametofüütsest isesobimatusest ei ole siin S-alleelid alati kodominantsed ja see muudabki asja keerulisemaks.
Nii gametofüütse kui ka sporofüütse isesobimatuse mehhanismi täielikuks mõistmiseks napib veel teadmisi. Näiteks pole täiesti selge, kuidas võetakse vastu ja antakse edasi signaale, mis aitavad tolmutoru(tera) ära tunda [8]. Alati ei tuntagi isesobimatut tolmutera ära, see sõltub partnerite ja nende suguorganite vanusest. Hilisfertiilsus (end-season-fertility) on nähtus, kus muidu isesobimatul liigil puhkevad vegetatsiooniperioodi lõpul viljastumisvõimelised õied [13]. Isesobimatust võib ületada ka üleküpsenud emaka viljastamisel värske õietolmuga, sest S-geeni kontrollitud produktide kontsentratsioon emaka koes on vähenenud [1]. Seega pakub loodus võimalusi ka seda üsna kindlalt toimivat barjääri parem ise kui ilma olukorras vältida.
Järgmises ajakirjanumbris tuleb juttu sellest, kuidas on õistaimedel sugu määratud, millal inimene taimede sugulisuse avastas ja selle mõtte omaks võttis.
1. Ascher, P. D.; Peloqiun, S. J. 1966. Effect of floral ageing on the growth of compatible and incompatible pollen tubes in Lilium longiflorum. Amer. J. Bot. 53: 99102.
2. Bannikova, V. P.; Khvedynich, O. A. 1982. Plant Embryology. Naukova Dumka, Kiev: 163. (in Russian)
3. Darwin, Charles 1977. The Different Forms on Plants of the Same Species. Murray, London: 352.
4. Eisikowitch, Dan.; Woodell, S. R. J. 1975. The effect of water on pollen germination in two species of Primula. Evolution 28: 692694.
5. F¿egri Knut, van der Pijl, Leendert 1982. The Principles of Pollination Ecology. Mir, Moskva: 377. (in Russian)
6. Hess, Dieter 1983. Die Blüte. Ulmer, Stuttgart.
7. Knuth, P. 18951905. Handbuch der blütenbiologie. IIII. Leipzig, Engelmann.
8. Matton, Daniel Philippe et al. 1994. Self-incompatibility: How plants avoid illegitimate offspring. Proc. Natl. Acad. Sci. USA 91 (3): 19921997.
9. McClure, Bruce A. et al. 1989. Style self-incompatibility gene products of Nicotiana alata are ribonucleases. Nature 342: 955957.
10. McClure, Bruce A. et al. 1990. Self-incompatibility in Nicotiana alata involves degradation of pollen rRNA. Nature 347: 557560.
11. Nasrallah, June B. et al. 1994. Signalling the arrest of pollen tube development in self-incompatible plants. Science 266 (12): 15051508.
12. Reier, Ülle 1995. Õigete äratundmisest õites. Schola Biotheoretica 21, 6675.
13. Richards, A. John. 1986. Plant Breeding Systems. London, Chapman & Hall, xiv+529.
Sõnaseletusi:
alleel (allele) üks kahest või mitmest alternatiivsest geenivariandist, mis asuvad populatsiooni isendite homoloogiliste kromosoomide samas lookuses ja toimivad sama tunnuse kujunemisele, tekitades selle eri vorme või avaldumisastmeid. Meioosis alleelid lahknevad eri rakkudesse, st. välistavad teineteist.
retsessiivsus (recessivity) ühe tunnuse (alleeli) varjuvus tunnusepaaris (alleelipaaris) heterosügootse genotüübi puhul; tunnuse (sh. defekti, haiguse) avaldumine ainult homosügootsuse (aa) korral. Retsessiivsus on dominantsuse vastandnähtus.
dominantsus (dominance) domineerimine, ühe tunnuse (alleeli) prevaleerumine tunnusepaaris (alleelipaaris) teise üle; heterosügootse genotüübi (Aa) avaldumine samamoodi dominantse homosügootse genotüübiga (AA); selliselt prevaleerivat tunnust (alleeli) nimetatakse dominantseks. Polüalleelsuse korral võib esineda astmeline dominantsus (A¹>A²>A³). Dominantsus ja retsessiivsus on ühe nähtuse eri poolused.
kodominantsus (codominance) tunnuste (alleelide) võrdväärne ja teineteisest sõltumatu avaldumine heterosügootse genotüübi puhul; mõlemast alleelist määratud tunnusevariandi üheaegne avaldumine heterosügootse indiviidi (raku) fenotüübis. Alleelide kodominantne avaldumine on tavaline molekulaarsete tunnuste, sh. rakupinna antigeenide (nt. vererühmade) puhul.
Geneetikamõistete seletusi võib leida aadressilt http://biomedicum.ut.ee/~martv/genolex.html#K
(Mart Viikmaa, Klassikalise geneetika leksikon)
Ülle Reier (1955) on lõpetanud Tartu ülikooli botaanikuna ja töötab praegu TÜ botaanika ja ökoloogia instituudi teadurina. Kaasa aidanud taimetundmise edendamisele (taimemääraja kaasautor, õppetöö ülikoolis). Taimede kojalisus on teda huvitanud juba mõnda aega.
| 

![[IN ENGLISH]](images/gb.gif)