Mitmesse riiki jaguneb elusloodus? IITõnu Ploompuu |
|
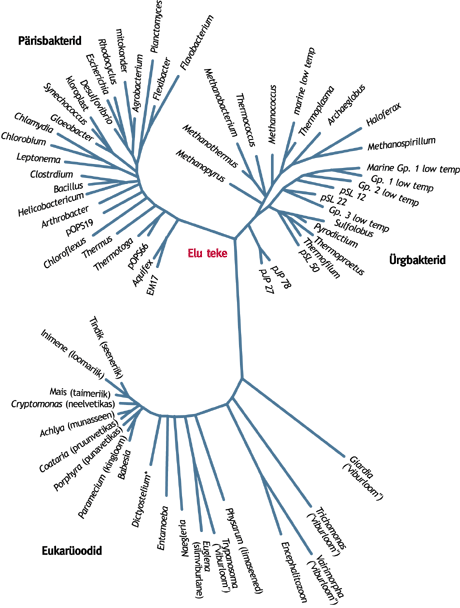
Eluslooduse süstematiseerimisel pole mitmeti defineeritav riik ainus võimalik suurim jaotus. Sõltuvalt sellest, kas lähtuda geenide arengust või organismide ja nende koosluste komplekssemaks muutumisest, võib leida veel teisigi suurjaotusi. Kolm domeeniViimasel ajal on hakatud rääkima kolmest elu algkodust - kolmest domeenist - lähtuvast evolutsioonipuust (joonis). Esimese haru moodustavad eukarüootsed (päristuumsed) organismid, kuhu kuuluvad kõik taimed, loomad, seened ja protistid. Teine haru on eubakterite (pärisbakterite) ehk teisi sõnu "vanade heade bakterite" päralt. Kolmandasse harusse kuuluvad suures osas alles viimasel paarikümnel aastal avastatud arhebakterid (ürgbakterid). Need on kemosünteesivad autotroofid, kes elavad sageli "ebanormaalsetes" tingimustes: kuumaveeallikates, tugevates soola- ja happelahustes, sügaval maapõues. Siiski on arhebakterite hulgas arvukalt ka tavalises merevees elavaid liike. Arhebakterite uurimine on alles pooleli ja võimalik, et kümmekonna aasta pärast ollakse nende levikust ja eluavaldustest veidi teisel arvamusel kui praegu. Nende kolme evolutsiooniharu diferentseerimine tugineb suurimatele põhimõttelistele erinevustele raku osade biokeemilises koostises ja ülesehituses. Domeen on oma olemuselt süstemaatiline ühik. Ta on suurem kui evolutsiooniharudel põhinev riik. Mitme riigi süsteemides on keeruline domeenisisene riigiline jaotumine levinud vaid päristuumsete puhul. Pärisbakteritel eristatakse fotosünteesivad bakterid mõnikord omaette alamriigiks, aga evolutsioonipuud vaadates tundub see olevat põhjendamatu - nende vastandumine on eluvormiline. Pärisbakterite plahvatuslik lahknemine fotosünteesi ja hingamise tekkeaegadel on andnud palju sarnaste omadustega tänini püsinud organismide rühmi. Nende jaotamise teeb eriti keerukaks sage geenide ülekanne kaugete bakteriliikide vahel. Ürgbakterite domeen jaotub seevastu küllaltki selgelt kolmeks (alam)riigiks. Domeenidesse saab rühmitada vaid uute mitme riigi süsteemide riike. Nelja ja viie riigi süsteemi puhul ei tule see kõne alla, sest need riigid on diferentseeritud teistel alustel kui domeenid. Sel puhul tuleks näiteks bakteririik jaotada kahe domeeni vahel, ülejäänud riigid aga saaks koondada ühte domeeni. Mitmesse riiki kuulub inimene?Täpne vastus küsimusele, mis riiki kuulub inimene, on loomariiki ja bakteririiki. Bakteririiki seetõttu, et üks oluline raku osa - mitokonder - tuleb tema evolutsioonilise päritolu tõttu arvata bakterite hulka. Päristuumsed organismid on kujunenud kahe organismi sümbioosi tulemusena. "Koos elama" on hakanud raku tuum (ürg-päristuumne) ja mitokonder (algselt hapnikku hingav bakter). Taim toetub kogunisti kolmele "jalale": ka kloroplasti (plastiidi) eellane oli iseseisev organism - tuntuim fotosüneesiv bakter sinivetikas. Seega ei saa päristuumseid organisme paigutada ainult ühele evolutsioonipuu oksale, vaid tuleb nad tunnistada kahte riiki kuuluvateks. Naljaga pooleks võiks öelda, et loomad ja seened kuuluvad korraga kahte, taimed kolme bakteriliiki. Kas õigesti süstematiseerimine on võimalik?Suurte organismirühmade süstemaatika toetub geenijärjestuste analüüsile. Sel puhul on oluline, milline geen vaatluse alla võtta. Kuid ka analüüsi jaoks sama väärtusega geenide põhjal võib saada erinevad evolutsioonipuud. Elu algpäevadel oli geenide liikumine organismide vahel tõenäoliselt väga vaba. Geenikaupa süstematiseerides kuuluks iga bakter ilmselt paljudesse alamriikidesse, mõnda natuke rohkem, teise vähem. On teada, et suurem osa päristuumsete geenidest sarnaneb rohkem ürgbakterite, veidi väiksem osa pärisbakterite omadega. Nii on "vana armas evolutsioonipuu" vaid ilus mudel, areng on toimunud pigem võrguna ja evolutsioonipuu asemel tuleks rääkida komplekssest eluvõrgust. Kui üks liik paigutada geenianalüüsile tuginedes mõnda riiki või domeeni, tuleb osa võrguharusid läbi lõigata. Nii jääb ikkagi kehtima olukord, kus osa organismidest jääb oma kuuluvuselt kahe riigi/domeeni vahele. Süstemaatika ei ole absoluutne tõde, vaid mudel organismide rühmitamiseks. Mudel on vajalik selgema ülevaate saamiseks. Evolutsioonipuud on pidevalt muutunud täpsemaks ja keerulisemaks. Liiga keeruliseks nad jällegi muutuda ei tohi, sest siis tuleb nende tundmaõppimiseks kasutada omakorda lihtsamaid mudeleid. Nelja riigi süsteemis on olnud evolutsioonipuule allumatud näiteks need "tegelased", kes elavad vahel taime, vahel loomana (silmviburlased). Geenijärjestustest lähtuvates süsteemides on sellisteks "hulkuvate" geenide-kromosoomide abil kujunenud organismid. Ja selliseid "hulguseid" tundub olevat enamikus organismides mitu. Elu neli komplekssustasetSegasust riikideks jagamise mudelites ja süstemaatikas üldse aitab vähendada mõistmine, et paljud keerukad tervikorganismid on tekkinud lihtsamate liitumise tagajärjel. Samblike puhul on see vana tõde, päristuumsete taoline kujunemine on veendumuseks saanud alles viimasel kümnendil. Võib eristada nelja organismi meenutavat elusolendite komplekssustaset. Iga ülenev koosneb madalama taseme ühikutest. Mida kõrgemaks muutub tase, seda nõrgemaks jääb seos tema koosteosade vahel ja hägusemaks taseme piirid. Esimene tase - eeleluline. Tervikuna sobivad sellele tasemele viirused ja plasmiidid ("elus aine"), rakkudest võiks siia kuuluvatena käsitleda kromosoome või nende tugevamini seotud lõike. Üleneva taseme osadena on need ühikud sageli eristamatult liitunud. Teine tase - eeltuumne. Bakterite kõrval on siin ka päristuumsete rakkude kromosoomidega organellid: tuum, plastiidid, mitokondrid. Võimalik, et ka raku tuum on kujunenud bakterite varajase sümbioosi tulemusena (Ploompuu 1999). Üleneva taseme osadena on nad hästi eristuvad, aga pole üldjuhul enam autonoomsed. Kolmas tase - päristuumne. Siia kuuluvad taimed, loomad ja seened. Nende kohta kehtib enamik vanu rakubioloogia reegleid (mitoos, meioos, palju eri organelle). Ülenevasse tasemesse kuuluvatena on nad üsna autonoomsed ning saavad enamasti ka üksi hakkama. Neljas tase - sümbiootiline. Siin on eri päristuumsete liikidest koosnevad liitorganismid: samblikud, mõnede kilptäide ja nende seljas elavate seente ning korallide ja nendes kasvatatavate vetikate terviklikud kooselud. Üsna sarnane sellele tasemele on ka inimese kooselu soolekepikesega või liblikõielistel mügarbakteritega - ainult et see toimub erinevate komplekssustasemete vahel. Sellised liitorganismid esinevad looduses küll enamasti tervikuna ja omavad selgeid terviklikkuse näitajaid, kuid tehistingimustes on neid lihtne lahus kasvatada nagu teisigi sümbiootilisi organisme. Siit samm edasi ja oleme juba ökosüsteemides, mis mõnest küljest vaadatuna võivad samuti tervikorganisme meenutada. Kokkuvõtteks võib öelda, et ühtset vastust küsimusele, mitmesse riiki jaguneb elusloodus, pole. Palju sõltub sellest, mida käsitletakse, mis suunast vaadatakse, milleks on riikideks jaotamise mudelit vaja. Võib rääkida nii süstemaatilisest, ökoloogilisest ja isegi didaktilisest riigist. Võib-olla oleks nende puhul õigem kasutada ka erinevaid väljendeid, nimetades ökoloogilisi riike näiteks elu(viisi)riikideks? Riigi kui süstemaatilise ühiku kohale või ökoloogilise riigi kõrvale on konkureeriva suurima süstemaatilise jaotusena kerkinud veel domeen. Miks ei võiks siis ka komplekssustase olla elusolendite mitmekesisuse üks võimalik suurim jaotusühik? KIRJANDUS Pace N. R. 1997. A Molecular View of Microbial diversity and
the biosphere. Science, 276 |
![]()